jueves, 22 de abril de 2010
GAMETOGÉNESIS.
GAMETOGÉNESIS.
Es el proceso mediante el cual los gametos de ambos sexos se diferencian y maduran hasta ser aptos para la fecundación. En los gametos masculinos el proceso se denomina espermatogénesis y en los femeninos ovulogénesis.
1. ESPERMATOGÉNESIS.
Comienza en la pubertad, alcanza un máximo y decae hasta la senectud. Es un proceso continuo que tiene lugar en el testículo y las estructuras anejas.
Los testículos constituidos por los tubos seminíferos con un intersticio en el que se encuentran las células de Leydig, sobre el cual hay una membrana basal que delimita el túbulo. En dicha membrana asientan las espermatogonias y las células de Sertoli que tienen tres funciones :
- soporte y liberación de los espermatozoides.
- función de barrera.
- producción de proteína específica (proteína fijadora de andrógenos - ABP).
Los espermatozoides pasan de la luz de túbulos seminíferos a la tete testis - epedídimo - conducto deferente - vesícula seminal - conducto eyaculador - uretra.
2. OVULOGÉNESIS.
Se produce en el ovario y comienza en la vida embrionaria. Las células germinativas dan lugar a las OVOGONIAS y éstas, en vida uterina, se transforman en ovocitos primarios que se rodean de células foliculares para formar los FOLÍCULOS PRIMORDIALES. En la 20ª semana se detienen en la profase de la meiosis I. En la vida fetal llegan a ser varios millones, pero por atresia se reducen a 400.000 en la pubertad.
Durante la vida reproductiva, 36 horas antes de la ovulación y coincidiendo con el pico de LH, el ovocito del folículo de Graaf reanuda la meiosis I, y se forma el PRIMER CORPÚSCULO POLAR ( que aparta los cromosomas sobrantes) y el OVOCITO II, que es captado por la trompa y con la mitad de cromosomas que el ovocito I.
(La meiosis II sólo se completa si el espermatozoide penetra en el interior del ovocito, se expulsa el SEGUNDO CORPÚSCULO POLAR y se fusionan los pronúcleos).
Antes de la ovulación, el cúmulo ovígero se desprende de la granulosa y queda flotando en el líquido folicular que se expulsa con la ovulación.
La fimbria ovárica de la trompa se contrae y acerca el pabellón tubárico a la zona donde se produce la ruptura folicular. Así, el cúmulo entra en la trompa (los cilios de las fimbrias son capaces de crear una corriente capaz de recoger partículas del saco de Douglas).
Se distinguen tres fases :
- paso de la ampolla tubárica (36 h - fecundación)
- paso de la unión ampolla-istmo en relación de la progesterona
- paso del istmo.
Este transporte se produce por la membrana, el movimiento ciliar y por la secreción.
VEA PROCESO EN:
http://www.youtube.com/watch?v=MkZHV74MpZw&feature=related
Es el proceso mediante el cual los gametos de ambos sexos se diferencian y maduran hasta ser aptos para la fecundación. En los gametos masculinos el proceso se denomina espermatogénesis y en los femeninos ovulogénesis.
1. ESPERMATOGÉNESIS.
Comienza en la pubertad, alcanza un máximo y decae hasta la senectud. Es un proceso continuo que tiene lugar en el testículo y las estructuras anejas.
Los testículos constituidos por los tubos seminíferos con un intersticio en el que se encuentran las células de Leydig, sobre el cual hay una membrana basal que delimita el túbulo. En dicha membrana asientan las espermatogonias y las células de Sertoli que tienen tres funciones :
- soporte y liberación de los espermatozoides.
- función de barrera.
- producción de proteína específica (proteína fijadora de andrógenos - ABP).
Los espermatozoides pasan de la luz de túbulos seminíferos a la tete testis - epedídimo - conducto deferente - vesícula seminal - conducto eyaculador - uretra.
2. OVULOGÉNESIS.
Se produce en el ovario y comienza en la vida embrionaria. Las células germinativas dan lugar a las OVOGONIAS y éstas, en vida uterina, se transforman en ovocitos primarios que se rodean de células foliculares para formar los FOLÍCULOS PRIMORDIALES. En la 20ª semana se detienen en la profase de la meiosis I. En la vida fetal llegan a ser varios millones, pero por atresia se reducen a 400.000 en la pubertad.
Durante la vida reproductiva, 36 horas antes de la ovulación y coincidiendo con el pico de LH, el ovocito del folículo de Graaf reanuda la meiosis I, y se forma el PRIMER CORPÚSCULO POLAR ( que aparta los cromosomas sobrantes) y el OVOCITO II, que es captado por la trompa y con la mitad de cromosomas que el ovocito I.
(La meiosis II sólo se completa si el espermatozoide penetra en el interior del ovocito, se expulsa el SEGUNDO CORPÚSCULO POLAR y se fusionan los pronúcleos).
Antes de la ovulación, el cúmulo ovígero se desprende de la granulosa y queda flotando en el líquido folicular que se expulsa con la ovulación.
La fimbria ovárica de la trompa se contrae y acerca el pabellón tubárico a la zona donde se produce la ruptura folicular. Así, el cúmulo entra en la trompa (los cilios de las fimbrias son capaces de crear una corriente capaz de recoger partículas del saco de Douglas).
Se distinguen tres fases :
- paso de la ampolla tubárica (36 h - fecundación)
- paso de la unión ampolla-istmo en relación de la progesterona
- paso del istmo.
Este transporte se produce por la membrana, el movimiento ciliar y por la secreción.
VEA PROCESO EN:
http://www.youtube.com/watch?v=MkZHV74MpZw&feature=related
El Ciclo Menstrual

El ciclo menstrual constituye el periodo durante el cual el organismo de la mujer se prepara para un eventual embarazo. Comienza el primer día de la menstruación y termina el primer día de la siguiente. En general, tiene una duración promedio de 28 días, pero se considera normal entre 21 y 35 días. Considerando siempre que el ciclo mestrual tiene una duración de 28 días como promedio.
Todas las mujeres normales nacen con alrededor de 300 mil óvulos en sus ovarios, que se encuentran en estado inmaduro. El inicio del desarrollo sexual durante la pubertad motiva la maduración del primer óvulo y, por lo tanto, es el comienzo del primer ciclo menstrual, situación que en promedio ocurre a la edad de 12 años y que se denomina menarquia.
Esta condición marca el inicio de la etapa fértil que se extiende hasta la última menstruación, conocida como menopausia. Durante este lapso madura un óvulo cada mes, llegando a repetirse el fenómeno alrededor de 500 veces durante toda la vida reproductiva.
El ciclo menstrual tiene dos fases, la folicular, que se extiende desde la menstruación hasta la ovulación y la lútea, que comprende el periodo postovulatorio hasta la siguiente menstruación.
Traducción
Traducción..
Una vez que el ARNm se encuentra en el citoplasma, es reconocido por el ribosoma mediante secuencias específicas (en bacterias) y por la caperuza (en eucariotas). En el ribosoma se lleva a cabo el proceso de traducción. En este momento cobra importancia el ARNt, que funciona como adaptador entre aminoácidos y ARNm.
¿Cómo reconocen los ARNt qué aminoácidos deben colocar para traducir una secuencia determinada?
Los ARNt tienen una región que se une a un aminoácido específico y otra que reconoce un triplete de nucleótidos en el ARNm (anticodón). La traducción comienza cuando el ribosoma reconoce ciertas secuencias en el extremo 5’ del ARNm (en bacterias) o la caperuza (en eucariotas) y se mueve a lo largo del mensajero hasta que encuentra el primer codón AUG, que codifica para metionina (o formil-met en bacterias). Este codón funciona como sitio de inicio. A medida que avanza la traducción, distintos ARNt se van uniendo al codón que le corresponda, se forma el enlace peptídico entre los aminoácidos, y por último se libera el ARNt “descargado”, quedando unido al ribosoma el último ARNt incorporado “cargando” con la cadena peptídica en crecimiento.
La transcripción ocurre en el núcleo, donde se eliminan los intrones del pre-ARNm y se crea un ARN maduro, que migra al citoplasma. Una vez que este se une a los ribosomas (formados por subunidades de ARNr y proteínas) y al ARN de transferencia, comienza el proceso de traducción.
¿Cuándo termina la traducción?
Los tres codones que no son reconocidos por ningún ARNt (es decir, que no codifican ningún aminoácido) funcionan como señales de terminación. De esta manera, cuando aparece uno de estos tripletes (UAA, UAG, UGA) la proteína recién formada se libera del ribosoma.
Es importante destacar que en principio el ribosoma podría leer tres “oraciones diferentes”, tener tres marcos de lectura en el mismo ARNm, ya que puede comenzar a leer los tripletes en una base, en la base siguiente o en la tercera base. Es decir, un marco de lectura es una secuencia ininterrumpida de tripletes que puede ser diferente de acuerdo con el nucléotido de inicio.
A partir de una misma secuencia de ARN es posible “leer” tres oraciones diferentes, lo que se traduce en tres proteínas diferentes. La secuencia será completamente diferente e incluso puede variar su longitud si, por ejemplo, aparece un codón de terminación (codón UAA)
Procesos postraducción
Una vez traducidas, las proteínas deben adoptar una estructura tridimensional adecuada. Esto se logra con la ayuda de otras proteínas denominadas chaperonas moleculares. Luego pueden ser modificadas mediante la unión de distintas moléculas como azúcares, nucleósidos, o fosfatos y dirigidas a lugares específicos de la célula (la membrana celular, el núcleo, etcétera) de acuerdo con su función.
Cuando una proteína cumplió su función o cuando no se plegó correctamente, es degradada. Las proteínas que van a ser degradadas son “marcadas” por la unión de una proteína llamada ubiquitina. Una vez seleccionadas, un complejo multiproteico (el proteosoma), degrada las proteínas en aminoácidos rompiendo los enlaces peptídicos.
Una vez que el ARNm se encuentra en el citoplasma, es reconocido por el ribosoma mediante secuencias específicas (en bacterias) y por la caperuza (en eucariotas). En el ribosoma se lleva a cabo el proceso de traducción. En este momento cobra importancia el ARNt, que funciona como adaptador entre aminoácidos y ARNm.
¿Cómo reconocen los ARNt qué aminoácidos deben colocar para traducir una secuencia determinada?
Los ARNt tienen una región que se une a un aminoácido específico y otra que reconoce un triplete de nucleótidos en el ARNm (anticodón). La traducción comienza cuando el ribosoma reconoce ciertas secuencias en el extremo 5’ del ARNm (en bacterias) o la caperuza (en eucariotas) y se mueve a lo largo del mensajero hasta que encuentra el primer codón AUG, que codifica para metionina (o formil-met en bacterias). Este codón funciona como sitio de inicio. A medida que avanza la traducción, distintos ARNt se van uniendo al codón que le corresponda, se forma el enlace peptídico entre los aminoácidos, y por último se libera el ARNt “descargado”, quedando unido al ribosoma el último ARNt incorporado “cargando” con la cadena peptídica en crecimiento.
La transcripción ocurre en el núcleo, donde se eliminan los intrones del pre-ARNm y se crea un ARN maduro, que migra al citoplasma. Una vez que este se une a los ribosomas (formados por subunidades de ARNr y proteínas) y al ARN de transferencia, comienza el proceso de traducción.
¿Cuándo termina la traducción?
Los tres codones que no son reconocidos por ningún ARNt (es decir, que no codifican ningún aminoácido) funcionan como señales de terminación. De esta manera, cuando aparece uno de estos tripletes (UAA, UAG, UGA) la proteína recién formada se libera del ribosoma.
Es importante destacar que en principio el ribosoma podría leer tres “oraciones diferentes”, tener tres marcos de lectura en el mismo ARNm, ya que puede comenzar a leer los tripletes en una base, en la base siguiente o en la tercera base. Es decir, un marco de lectura es una secuencia ininterrumpida de tripletes que puede ser diferente de acuerdo con el nucléotido de inicio.
A partir de una misma secuencia de ARN es posible “leer” tres oraciones diferentes, lo que se traduce en tres proteínas diferentes. La secuencia será completamente diferente e incluso puede variar su longitud si, por ejemplo, aparece un codón de terminación (codón UAA)
Procesos postraducción
Una vez traducidas, las proteínas deben adoptar una estructura tridimensional adecuada. Esto se logra con la ayuda de otras proteínas denominadas chaperonas moleculares. Luego pueden ser modificadas mediante la unión de distintas moléculas como azúcares, nucleósidos, o fosfatos y dirigidas a lugares específicos de la célula (la membrana celular, el núcleo, etcétera) de acuerdo con su función.
Cuando una proteína cumplió su función o cuando no se plegó correctamente, es degradada. Las proteínas que van a ser degradadas son “marcadas” por la unión de una proteína llamada ubiquitina. Una vez seleccionadas, un complejo multiproteico (el proteosoma), degrada las proteínas en aminoácidos rompiendo los enlaces peptídicos.
Transcripción
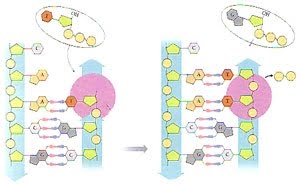
Una vez que se conforman las dos cadenas nuevas de ADN, lo que sigue es pasar la información contenida en estas cadenas a una cadena de ARN, proceso que se conoce como transcripción. Aquí la enzima responsable es la ARN polimerasa, la cual se une a una secuencia específica en el ADN denominada promotor y sintetiza ARN a partir de ADN.
En la transcripción, la información codificada en un polímero formado por la combinación de 4 nucleótidos (ADN) se convierte en otro polímero cuyas unidades también son 4 nucleótidos (ARN). El ácido ribonucleico es similar al ADN (por eso el proceso se denomina transcripción), pero poseen algunas diferencias, como mostramos en la figura
La transcripción de genes puede dar lugar a ARN mensajero (ARNm, molécula que sirve como molde de la traducción), ARN ribosomal (ARNr, que forma parte de los ribosomas, un complejo compuesto por proteínas y ARNr donde se realiza el proceso de traducción) o ARN de transferencia (ARNt, moléculas que funcionan como adaptadores en el proceso de traducción).
Fenómenos postranscripción
Si bien estos pasos básicos son los mismos para la mayoría de los organismos, hay diferencias entre los distintos dominios de seres vivos (Bacteria, Arquea y Eucaria) tanto en la replicación como en la transcripción y en la traducción.
En organismos procariontes, el ARNm se une a los ribosomas y puede ser traducido tal y como es liberado de la ARN polimerasa, ya que se encuentra en el citoplasma celular y no sufre ninguna modificación. Incluso, puede ser traducido a medida que es transcripto.
En los eucariontes, sin embargo, la traducción y transcripción ocurren en forma separada. La transcripción ocurre en el núcleo y la traducción, en el citoplasma, puede ocurrir minutos, horas o incluso días más tarde. Antes de salir del núcleo para ser traducido, el ARNm sufre dos modificaciones, por lo que es llamado pre-ARNm.
La primera de ellas es el procesamiento por corte y empalme (splicing, en inglés), en el cual se eliminan algunos secuencias no codificantes (o intrones) y se unen las secuencias codificantes (exones). Una molécula de ARNm puede llegar a tener hasta 70 intrones, que pueden llegar a variar de tamaño entre 80 y 10.000 nucleótidos. La segunda modificación ocurre en los extremos: al extremo 5’ se le une una caperuza (compuesta por guanina metilada) y al extremo 3’ se agrega una “cola” de poliadenina o poliA. Luego de todas estas modificaciones, tenemos un ARN maduro.
Replicación del ADN
En este apartado describimos con más detalle los procesos de replicación, transcripción y traducción, introducidos en el apartado anterior.
La transmisión de información implica que el ADN es capaz de duplicarse de manera de obtener dos moléculas iguales a partir de la molécula inicial. Este proceso se llama replicación.
Luego del descubrimiento de la estructura del ADN, en 1957, dos biólogos moleculares americanos, Matthew Stanley Meselson y Frank Stahl demostraron que este se replica de una manera semiconservativa, es decir que la nueva cadena se sintetiza utilizando una de las hebras preexistentes como molde. Las moléculas de ADN “hijas” están formadas por una cadena nueva y una original que sirve como molde. Con nitrógeno 15 (un isótopo radiactivo), ya que el nitrógeno es necesario para la síntesis de las bases que componen el ADN, y usando sucesivas generaciones de bacterias Escherichia coli, estos científicos mostraron que cuando el ADN se duplica, cada una de sus cadenas pasa a las células hijas sin cambiar y actúan de molde o patrón para formar una segunda hebra y completar así las dos doble cadenas.
Para que esto ocurra, la célula debe “abrir” la doble cadena de ADN en una secuencia específica denominada origen de replicación(en bacterias) o secuencia de replicación autónoma (en eucariotas) y copiar cada cadena.
En la replicación participan varias enzimas. Las ADN polimerasas sintetizan una nueva cadena de ADN. Para esto utilizan como molde una de las hebras y un segmento corto de ADN, al que se le agregan los nuevos nucleótidos. Este segmento funciona como cebador (primer, en inglés). La ADN polimerasa agrega nucleótidos al extremo 3’ de la cadena en crecimiento.
martes, 20 de abril de 2010
Mitocondrias
Son organelos citoplasmáticos membranosos característicos de las células eucarióticas. Se habrían originado, al igual que los cloroplastos, en momentos tempranos de la evolución a partir de la endocitosis de bacterias fotosintéticas (teoría endosimbiótica). Las mitocondrias poseen una gran importancia, ya que en ellas se realizan una serie de reacciones de óxido-reducción que permiten el sustento energético de la célula. De esta manera, y de forma general, células que realizan un mayor gasto de energía poseerán una mayor cantidad de mitocondrias. Su dimension varia entre 1 a 10 u.
Estructura
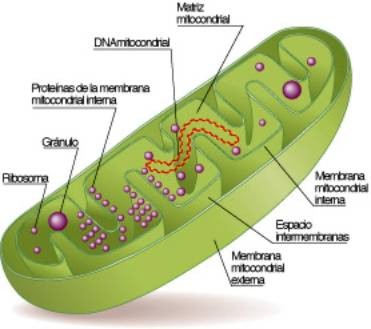
La microscopia de luz muestra a las mitocondrias nítidamente teñidas de manera uniforme ; pero para dilucidar su verdadera estructura debemos de valernos de la microscopia electronica con mayor poder resolutivo. Demostrando entonces que la mitocondria consta de 2 membranas paralelas separadas por un espacio estrecho(membrana externa e interna en razón a su ubicación ) o interestructural .Este espacio esta ocupado por una Sustancia homogénea denominada matriz mitocondrial que es de menor opacidad a los rayos electrónicos ,contienen enzimas del ciclo del acido citrico .la membrana interna emite hacia el interior de la mitocondria evaginaciones laminares a manera de tabiques incompletos que no llegan a segmentar el espacio interestructural .Estas prolongaciones se denominan crestas mitocondriales.
Variantes de las crestas
Generalmente las crestas son laminares y perpendiculares al eje mayor de la mitocondria pero existen crestas irregulares en forma de dedo tubulares ,o paralelas al eje como en los espermatogonios humanos a veces encontramos crestas angulares ;también es variable el numero de crestas, asi existen mitocondrias con crestas escasas y cortas o por el contrario hay casos en los que las crestas son supernumerarias y casi no dejan espacio para la matriz intramitocondrial .
Suscribirse a:
Entradas (Atom)
